Sensibilità ambientale e varianza genica del trasportatore della serotonina: dai ratti agli esseri umani
7th November 2020 - By Judith R Homberg
Sull’autore/autrice
Judith Homberg è professoressa di Neuroscienze Traslazionali presso il Donders Institute for Brain, Cognition, and Behaviour a Nijmegen, Paesi Bassi. Il suo gruppo di ricerca preclinico si concentra sulla comprensione delle differenze individuali nel comportamento in relazione alla vulnerabilità e alla resilienza ai disturbi legati al neurosviluppo e allo stress.
Riassunto
La sensibilità ambientale è osservata sia negli animali che negli esseri umani, suggerendo che è ha una solida base biologica. Un fattore biologico che aumenta la sensibilità ambientale è il gene trasportatore di serotonina. Questo studio ha dimostrato che la sensibilità ambientale è associata a substrati neurali simili negli esseri umani e negli animali con regolazione a basso down del trasportatore di serotonina ereditaria.
Informazioni di base
Si pensa che la sensibilità ambientale abbia le sue radici nell’evoluzione. In effetti, è pertinente per la stabilità complessiva di molte popolazioni animali, comprese le popolazioni umane, che ci sono individui che sono audaci e poco influenzati dall’ambiente, nonché individui cauti e altamente sensibili all’ambiente. Tale base evolutiva suggerisce che le differenze individuali nella sensibilità ambientale hanno una solida base biologica.
Differenze genetiche nella sensibilità
Una varianza genica esemplare che conferisce sensibilità ambientale è la regione polimorfica legata al trasportatore di serotonina (5-HTTLPR), che descrive una sezione sul DNA che viene fornita in una versione breve o lunga, indicata come varianti alleliche La variante allelica corta, rispetto alla variante allelica lunga, di questa varianza genica promotore è associata a una ridotta espressione del trasportatore di serotonina (5-HTT) e ad una maggiore sensibilità ambientale.
Questa varianza genica si trova negli esseri umani e nei primati non umani, ma non in altre specie animali. A causa delle differenze di specie nella regione promotore del gene 5-HTT, i tentativi di ingegneria genetica di indurre il 5-HTTLPR in altre specie animali fallirono. Tuttavia, può essere modellato nei roditori attraverso il knockout 5-HTT, quindi l’inattivazione del gene 5-HTT.
Mentre un knockout genico non è una copia esatta di un polimorfismo promotore, c’è una sorprendente sovrapposizione negli effetti comportamentali del polimorfismo negli esseri umani e nei primati non umani e nel knockout genico nei roditori.
Così, gli esseri umani, primati non umani e topi knockout trasportatore di serotonina tutti dimostrano una maggiore cautela e ansia, e una maggiore responsabilità a entrambi gli stimoli ambientali avversi e gratificanti.
Nell’uomo queste possono essere immagini di volti tristi e felici, e nello stress degli animali (ad esempio la sconfitta sociale) e la ricompensa (ad esempio saccarosio, arricchimento ambientale) [1, 2]. Ciò suggerisce che la funzione del trasportatore di serotonina è altamente conservata e che i suoi effetti sul comportamento sono mediati attraverso meccanismi neurali simili (cioè, gli effetti genetici influenzano il comportamento attraverso specifici meccanismi neurali). Tuttavia, questo non è stato testato empiricamente.
Obiettivi di studio
Lo scopo del nostro studio [2] era quello di esaminare se la sensibilità ambientale negli esseri umani che trasportano il polimorfismo promotore del trasportatore di serotonina e i ratti privi del trasportatore di serotonina sono mediati attraverso neurocircufuitries simili.
Come è stato condotto lo studio umano
Ci siamo concentrati sulla sensibilità a uno stimolo negativo (minaccia), quindi su un lato della sensibilità ambientale, perché la sensibilità alle minacce può essere testata sia negli esseri umani che nei ratti. Abbiamo testato volontari umani sani (N.104) in un paradigma di condizionamento della paura probabilistica (cioè, un esperimento in cui i partecipanti hanno imparato ad accoppiare uno stimolo con un’esperienza negativa), che è stato condotto in uno scanner cerebrale (MRe) e combinato con misurazioni della frequenza cardiaca utilizzando un ossimetro.
Gli esseri umani hanno ricevuto scosse elettriche attraverso un indice delle dita, mentre guardavano semplici segnali presentati su uno schermo. Uno dei segnali prevedeva gli shock con una probabilità del 30%. Abbiamo determinato in modo specifico la risposta neurale fisiologica e associata ai segnali di previsione delle minacce in assenza della minaccia (shock elettrico) stessa.
Principali risultati
Abbiamo scoperto che in risposta alle minacce gli individui che trasportavano la variante allelica corta 5-HTTLPR mostravano una maggiore paura della bradcardia (una riduzione della frequenza cardiaca) e che questo è stato mediato da una maggiore connettività tra l’amigdala (situata in profondità nel cervello e l’emozione di “segnalazione”) e il grigio periaqueductale (situato nel gambo cerebrale e mediando le risposte autonomiche come la frequenza cardiaca).
Come è stato condotto lo studio sugli animali
Per determinare se i ratti da knockout 5-HTT avrebbero display simili correlazioni fisiologiche e neurali, abbiamo sottoposto i ratti knockout e control 5-HTT a un paradigma di condizionamento della paura e estinzione.
Durante la paura condizionare i toni udici sono stati associati con footshocks. Durante l’estinzione della paura i toni sono stati presentati senza footshock e sono state misurate le risposte comportamentali e fisiologiche.
Principali risultati
Abbiamo osservato che i ratti da knockout 5-HTT mostravano un aumento del congelamento (nessun movimento tranne la respirazione) e temevano la bradicardia, come nei portatori di alleli corti 5-HTTLPR umani. Il congelamento può riflettere ‘pausa-to-check’ come si osserva in genere in individui prudenti.
Per studiare se il congelamento e la paura della bradicardia fossero in ratti da knockout 5-HTT mediati anche da una connettività alterata tra l’amigdala e il grigio periaqueductale, abbiamo condotto immunostainings ex vivo, concentrandoci sui neuroni positivi della somatostatina GABAergic nell’amigdala noti per proiettare sul grigio periaqueductale.
L’attività di questi neuroni è stata aumentata nel knockout 5-HTT e ha scoperto di mediare in modo significativo l’associazione tra il gene 5-HTT e il congelamento [3].
Conclusioni generali
Questo studio dimostra che le somiglianze comportamentali e fisiologiche tra i portatori di alleli corti 5-HTTLPR umani e i roditori knockout 5-HTT hanno correlazioni neurali simili, rendendo provvisorio l’uso dei roditori per comprendere ulteriormente la base neurobiologica della sensibilità ambientale.
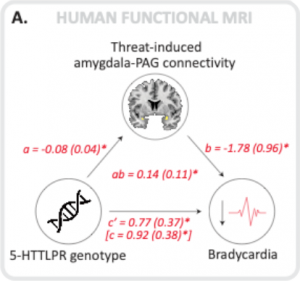
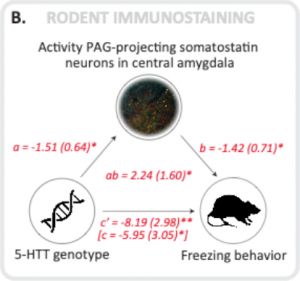
A mediation analysis shows that both in humans and in rats the connectivity between the amygdala and the periaqueductal gray (PAG) significantly mediates the association between inherited serotonin transporter (5-HTT) gene variance and fear bradycardia (humans) and freezing rats.
Bibliografia
- Homberg, J.R. and K.P. Lesch, Looking on the bright side of serotonin transporter gene variation. Biol Psychiatry, 2011. 69(6): p. 513-9.
- Homberg, J., et al., Sensory processing sensitivity and serotonin gene variance: Insights into mechanisms shaping environmental sensitivity. . Neurosci Biobehav Rev, 2016. 71: p. 472-483.
- Schipper, P., et al., The association between serotonin transporter availability and the neural correlates of fear bradycardia. Proc Natl Acad Sci U S A, 2019. 116(51): p. 25941-25947.